


Le monde qui nous entoure est rempli de toutes sortes d’informations que notre cerveau traite en permanence. Il reçoit ces informations par l'intermédiaire des organes des sens, dont chacun est responsable de sa part de signaux : les yeux (vision), la langue (goût), le nez (odorat), la peau (toucher), l'appareil vestibulaire (équilibre, position dans l'espace et sens de la poids) et les oreilles (son). En combinant les signaux de tous ces organes, notre cerveau peut se forger une image précise de notre environnement. Mais nous ne connaissons pas tous les aspects du traitement des signaux externes. L'un de ces secrets est le mécanisme permettant de localiser la source des sons.
Des scientifiques du Laboratoire de neuro-ingénierie de la parole et de l'audition (Institut de technologie du New Jersey) ont proposé un nouveau modèle du processus neuronal de localisation du son. Quels processus exacts se produisent dans le cerveau lors de la perception du son, comment notre cerveau comprend la position de la source sonore et comment cette recherche peut aider à lutter contre les défauts auditifs. Nous l’apprenons grâce au rapport du groupe de recherche. Aller.
Base de recherche
Les informations que notre cerveau reçoit de nos sens diffèrent les unes des autres, tant en termes de source que de traitement. Certains signaux apparaissent immédiatement à notre cerveau comme des informations précises, tandis que d’autres nécessitent des processus informatiques supplémentaires. En gros, on ressent immédiatement un toucher, mais quand on entend un son, encore faut-il trouver d'où il vient.
La base pour localiser les sons dans le plan horizontal est interaural* décalage horaire (ITD de décalage horaire interauriculaire) les sons parviennent aux oreilles de l'auditeur.
Base interaurale* - distance entre les oreilles.
Il existe une zone spécifique dans le cerveau (l’olive médiale supérieure ou MSO) qui est responsable de ce processus. Au moment où le signal sonore est reçu dans le MVO, les différences de temps interaurales sont converties en vitesse de réaction des neurones. La forme des courbes de vitesse de sortie MBO en fonction de l'ITD ressemble à la forme de la fonction de corrélation croisée des signaux d'entrée pour chaque oreille.
La manière dont les informations sont traitées et interprétées dans le MBO reste pas tout à fait claire, c'est pourquoi il existe plusieurs théories très contradictoires. La théorie la plus célèbre et en fait classique de la localisation sonore est le modèle de Jeffress (Lloyd A. Jeffress). C'est basé sur ligne marquée* neurones détecteurs sensibles à la synchronisation binaurale des entrées neuronales de chaque oreille, chaque neurone étant sensible au maximum à une certaine quantité d'ITD (1А).
Principe de la ligne marquée* est une hypothèse qui explique comment différents nerfs, qui utilisent tous les mêmes principes physiologiques pour transmettre des impulsions le long de leurs axones, sont capables de générer des sensations différentes. Des nerfs structurellement similaires peuvent générer des perceptions sensorielles différentes s’ils sont connectés à des neurones uniques du système nerveux central capables de décoder des signaux nerveux similaires de différentes manières.
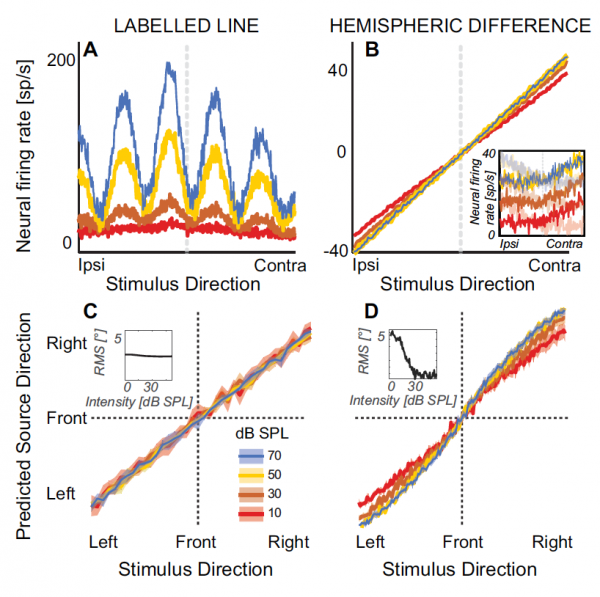
Image #1
Ce modèle est informatiquement similaire au codage neuronal, basé sur des corrélations croisées sans contrainte de sons atteignant les deux oreilles.
Il existe également un modèle qui suggère que la localisation sonore peut être modélisée sur la base des différences dans la vitesse de réponse de certaines populations de neurones de différents hémisphères du cerveau, c'est-à-dire modèle d'asymétrie interhémisphérique (1V).
Jusqu'à présent, il était difficile de déterminer sans ambiguïté laquelle des deux théories (modèles) était correcte, étant donné que chacune d'entre elles prédit des dépendances différentes entre la localisation du son et l'intensité sonore.
Dans l'étude que nous examinons aujourd'hui, les chercheurs ont décidé de combiner les deux modèles pour comprendre si la perception des sons est basée sur un codage neuronal ou sur des différences dans la réponse de populations neuronales individuelles. Plusieurs expériences ont été menées auxquelles ont participé des personnes âgées de 18 à 27 ans (5 femmes et 7 hommes). L'audiométrie des participants (une mesure de l'acuité auditive) était de 25 dB ou plus entre 250 et 8000 XNUMX Hz. Le participant aux expériences a été placé dans une pièce insonorisée, dans laquelle était placé un équipement spécial, calibré avec une grande précision. Les participants devaient, après avoir entendu un signal sonore, indiquer la direction d'où il venait.
Résultats de l'étude
Pour évaluer la dépendance latéralisation* activité cérébrale à partir de l'intensité sonore en réponse aux neurones marqués, des données sur la vitesse de réaction des neurones dans le noyau laminaire du cerveau de la chouette effraie ont été utilisées.
Latéralité* - asymétrie des moitiés gauche et droite du corps.
Pour évaluer la dépendance de la latéralisation de l'activité cérébrale sur la vitesse de réaction de certaines populations de neurones, des données provenant de l'activité du colliculus inférieur du cerveau du singe rhésus ont été utilisées, après quoi les différences de vitesse des neurones de différents hémisphères ont également été calculées. .
Le modèle de ligne marquée des neurones détecteurs prédit qu'à mesure que l'intensité sonore diminue, la latéralité de la source perçue convergera vers des valeurs moyennes similaires au rapport entre les sons faibles et les sons forts (1S).
Le modèle d'asymétrie hémisphérique, quant à lui, suggère qu'à mesure que l'intensité sonore diminue jusqu'à des niveaux proches du seuil, la latéralité perçue se déplacera vers la ligne médiane (1D).
À une intensité sonore globale plus élevée, la latéralisation devrait être invariante en intensité (encarts dans 1S и 1D).
Par conséquent, analyser comment l'intensité sonore affecte la direction perçue du son nous permet de déterminer avec précision la nature des processus qui se produisent à ce moment-là - neurones de la même zone générale ou neurones de différents hémisphères.
De toute évidence, la capacité d'une personne à distinguer les ITD peut varier en fonction de l'intensité sonore. Cependant, les scientifiques affirment qu'il est difficile d'interpréter les résultats antérieurs liant la sensibilité à l'ITD et le jugement des auditeurs sur la direction de la source sonore en fonction de l'intensité sonore. Certaines études affirment que lorsque l’intensité sonore atteint un seuil limite, la latéralité perçue de la source diminue. D'autres études suggèrent qu'il n'y a aucun effet de l'intensité sur la perception.
En d’autres termes, les scientifiques laissent entendre « gentiment » qu’il existe peu d’informations dans la littérature concernant la relation entre l’ITD, l’intensité sonore et la détermination de la direction de sa source. Il existe des théories qui existent comme des sortes d'axiomes, généralement acceptées par la communauté scientifique. Par conséquent, il a été décidé de tester en détail toutes les théories, modèles et mécanismes possibles de perception auditive dans la pratique.
La première expérience était basée sur un paradigme psychophysique qui permettait d'étudier la latéralisation basée sur l'ITD en fonction de l'intensité sonore dans un groupe de dix participants entendants normaux.
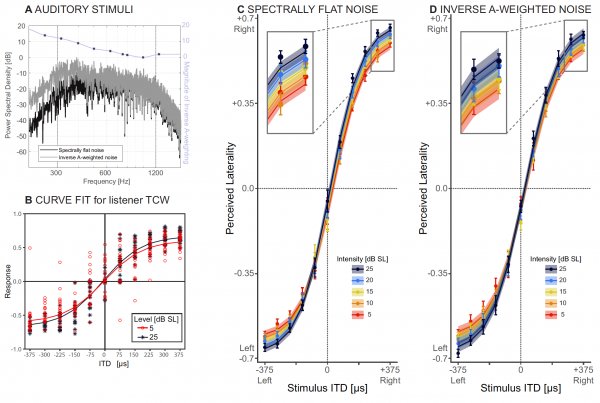
Image #2
Les sources sonores ont été spécifiquement réglées pour couvrir la majeure partie de la plage de fréquences dans laquelle les humains sont capables de détecter l'ITD, c'est-à-dire de 300 à 1200 Hz (2А).
A chaque essai, l'auditeur devait indiquer la latéralité perçue, mesurée en fonction du niveau de sensation, sur une plage de valeurs ITD allant de 375 à 375 ms. Pour déterminer l'effet de l'intensité sonore, un modèle à effets mixtes non linéaires (NMLE) a été utilisé, incluant à la fois une intensité sonore fixe et aléatoire.
Calendrier 2V démontre une latéralisation estimée avec un bruit spectralement plat à deux intensités sonores pour un auditeur représentatif. Et le calendrier 2S montre les données brutes (cercles) et le modèle NMLE ajusté (lignes) de tous les auditeurs.
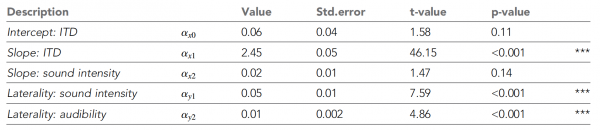
Tableau n ° 1
Le tableau ci-dessus montre tous les paramètres NLME. On peut voir que la latéralité perçue augmente avec l’augmentation de l’ITD, comme s’y attendaient les scientifiques. À mesure que l'intensité sonore diminuait, la perception se déplaçait de plus en plus vers la ligne médiane (encadré dans le graphique 2C).
Ces tendances ont été corroborées par le modèle NLME, qui a montré des effets significatifs de l'ITD et de l'intensité sonore sur le degré maximal de latéralité, confortant ainsi le modèle des différences interhémisphériques.
De plus, les seuils audiométriques moyens pour les sons purs avaient peu d’effet sur la latéralité perçue. Mais l'intensité sonore n'a pas affecté de manière significative les indicateurs des fonctions psychométriques.
L'objectif principal de la deuxième expérience était de déterminer comment les résultats obtenus dans l'expérience précédente changeraient en tenant compte des caractéristiques spectrales des stimuli (sons). La nécessité de tester le bruit spectralement plat à faible intensité sonore est que certaines parties du spectre peuvent ne pas être audibles, ce qui peut affecter la détermination de la direction du son. Par conséquent, les résultats de la première expérience peuvent être confondus avec le fait que la largeur de la partie audible du spectre peut diminuer avec la diminution de l'intensité sonore.
Par conséquent, il a été décidé de mener une autre expérience, mais en utilisant l'inverse Pondération A* bruit
Pesée A* appliqué aux niveaux sonores pour prendre en compte le volume relatif perçu par l’oreille humaine, puisque l’oreille est moins sensible aux basses fréquences sonores. La pondération A est mise en œuvre en ajoutant arithmétiquement un tableau de valeurs répertoriées en bandes d'octave aux niveaux de pression acoustique mesurés en dB.
Sur le graphique 2D montre les données brutes (cercles) et les données ajustées au modèle NMLE (lignes) de tous les participants à l'expérience.
L'analyse des données a montré que lorsque toutes les parties du son sont à peu près également audibles (à la fois dans le premier et dans le deuxième essai), la latéralité perçue et la pente dans le graphique expliquant le changement de latéralité avec l'ITD diminuent avec la diminution de l'intensité sonore.
Ainsi, les résultats de la deuxième expérience ont confirmé les résultats de la première. Autrement dit, dans la pratique, il a été démontré que le modèle proposé en 1948 par Jeffress n'est pas correct.
Il s'avère que la localisation du son se détériore à mesure que l'intensité sonore diminue, et Jeffress pensait que les sons sont perçus et traités par les humains de la même manière, quelle que soit leur intensité.
Pour une connaissance plus détaillée des nuances de l'étude, je recommande de regarder .
Le final
Des hypothèses théoriques et des expériences pratiques les confirmant ont montré que les neurones cérébraux des mammifères sont activés à des rythmes différents selon la direction du signal sonore. Le cerveau compare ensuite ces vitesses entre tous les neurones impliqués dans le processus pour construire dynamiquement une carte de l'environnement sonore.
Le modèle de Jeffresson n'est en fait pas faux à 100 %, puisqu'il peut être utilisé pour décrire parfaitement la localisation de la source sonore chez les effraies des clochers. Oui, pour les effraies des clochers, l’intensité du son n’a pas d’importance ; dans tous les cas, ce sont elles qui détermineront la position de sa source. Cependant, ce modèle ne fonctionne pas avec les singes rhésus, comme l’ont montré des expériences antérieures. Par conséquent, ce modèle de Jeffresson ne peut pas décrire la localisation des sons pour tous les êtres vivants.
Des expériences avec des participants humains ont confirmé une fois de plus que la localisation du son se produit différemment selon les organismes. De nombreux participants n'ont pas pu déterminer correctement la position de la source des signaux sonores en raison de la faible intensité des sons.
Les scientifiques pensent que leurs travaux montrent certaines similitudes entre la façon dont nous voyons et la façon dont nous entendons. Les deux processus sont associés à la vitesse des neurones dans différentes parties du cerveau, ainsi qu'à l'évaluation de cette différence pour déterminer à la fois la position des objets que nous voyons dans l'espace et la position de la source du son que nous entendons.
À l’avenir, les chercheurs mèneront une série d’expériences pour examiner plus en détail le lien entre l’audition et la vision humaines, ce qui nous permettra de mieux comprendre comment notre cerveau construit dynamiquement une carte du monde qui nous entoure.
Merci de votre attention, restez curieux et bonne semaine à tous ! 🙂
Merci de rester avec nous. Vous aimez nos articles ? Vous voulez voir du contenu plus intéressant ? Soutenez-nous en passant une commande ou en recommandant à vos amis, , 30 % de réduction pour les utilisateurs Habr sur un analogue unique de serveurs d'entrée de gamme, que nous avons inventé pour vous : (disponible avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Dell R730xd 2 fois moins cher ? Ici seulement aux Pays-Bas! Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - à partir de 99$ ! En savoir plus
Source: habr.com
