


De wereld om ons heen is gevuld met allerlei informatie die onze hersenen continu verwerken. Hij ontvangt deze informatie via zintuigen, die elk verantwoordelijk zijn voor hun deel van de signalen: ogen (visie), tong (smaak), neus (geur), huid (tastzin), vestibulaire apparaten (evenwicht, positie in de ruimte en gevoel voor gevoel). gewicht) en oren (geluid). Door signalen van al deze organen te combineren, kunnen onze hersenen een nauwkeurig beeld van onze omgeving opbouwen. Maar niet alle aspecten van het verwerken van externe signalen zijn bij ons bekend. Een van deze geheimen is het mechanisme voor het lokaliseren van de geluidsbron.
Wetenschappers van het Laboratory of Neuroengineering of Speech and Hearing (New Jersey Institute of Technology) hebben een nieuw model voorgesteld van het neurale proces van geluidslokalisatie. Welke processen plaatsvinden er precies in de hersenen tijdens de perceptie van geluid, hoe ons brein de positie van de geluidsbron begrijpt en hoe dit onderzoek kan helpen in de strijd tegen gehoorafwijkingen. Dat leren we uit het rapport van de onderzoeksgroep. Gaan.
Onderzoeksbasis
De informatie die onze hersenen van onze zintuigen ontvangen, verschilt van elkaar, zowel qua bron als qua verwerking. Sommige signalen verschijnen onmiddellijk voor onze hersenen als nauwkeurige informatie, terwijl andere aanvullende rekenprocessen vereisen. Grofweg voelen we een aanraking meteen, maar als we een geluid horen, moeten we nog zoeken waar het vandaan komt.
De basis voor het lokaliseren van geluiden in het horizontale vlak is interauraal* tijdsverschil (ITD van interauraal tijdsverschil) geluiden die de oren van de luisteraar bereiken.
Interaurale basis* - afstand tussen de oren.
Er is een specifiek gebied in de hersenen (de mediale superieure olijf of MSO) dat verantwoordelijk is voor dit proces. Op het moment dat het geluidssignaal in de MVO wordt ontvangen, worden interaurale tijdsverschillen omgezet in de reactiesnelheid van neuronen. De vorm van de MBO-uitgangssnelheidscurven als functie van ITD lijkt op de vorm van de kruiscorrelatiefunctie van de ingangssignalen voor elk oor.
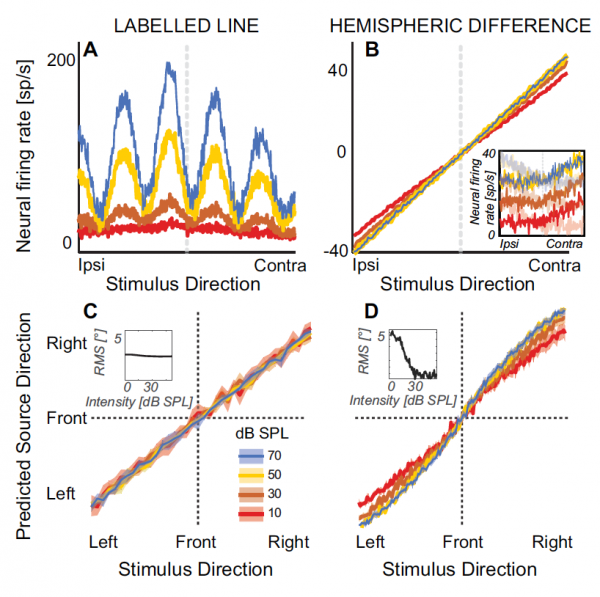
Hoe informatie in het mbo wordt verwerkt en geïnterpreteerd, blijft niet geheel duidelijk. Daarom bestaan er verschillende zeer tegenstrijdige theorieën. De bekendste en feitelijk klassieke theorie van geluidslokalisatie is het Jeffress-model (Lloyd A. Jeffres). Het is gebaseerd op gemarkeerde lijn* detectorneuronen die gevoelig zijn voor binaurale synchronisatie van neurale input van elk oor, waarbij elk neuron maximaal gevoelig is voor een bepaalde hoeveelheid ITD (1А).
Gemarkeerd lijnprincipe* is een hypothese die verklaart hoe verschillende zenuwen, die allemaal dezelfde fysiologische principes gebruiken bij het overbrengen van impulsen langs hun axonen, verschillende sensaties kunnen genereren. Structureel vergelijkbare zenuwen kunnen verschillende zintuiglijke waarnemingen genereren als ze verbonden zijn met unieke neuronen in het centrale zenuwstelsel die in staat zijn vergelijkbare zenuwsignalen op verschillende manieren te decoderen.
Afbeelding #1
Dit model is computationeel vergelijkbaar met neurale codering, gebaseerd op onbeperkte kruiscorrelaties van geluiden die beide oren bereiken.
Er is ook een model dat suggereert dat geluidslokalisatie kan worden gemodelleerd op basis van verschillen in de reactiesnelheid van bepaalde populaties neuronen uit verschillende hersenhelften, d.w.z. model van interhemisferische asymmetrie (1V).
Tot nu toe was het moeilijk om ondubbelzinnig te zeggen welke van de twee theorieën (modellen) juist is, aangezien elk van hen verschillende afhankelijkheden van geluidslokalisatie en geluidsintensiteit voorspelt.
In het onderzoek waar we vandaag naar kijken, besloten de onderzoekers beide modellen te combineren om te begrijpen of de perceptie van geluiden gebaseerd is op neurale codering of op verschillen in de reactie van individuele neurale populaties. Er zijn verschillende experimenten uitgevoerd waaraan mensen van 18 tot 27 jaar (5 vrouwen en 7 mannen) deelnamen. De audiometrie van de deelnemers (een meting van de gehoorscherpte) was 25 dB of hoger tussen 250 en 8000 Hz. De deelnemer aan de experimenten werd in een geluiddichte ruimte geplaatst, waarin speciale apparatuur werd geplaatst, gekalibreerd met hoge nauwkeurigheid. Deelnemers moesten bij het horen van een geluidssignaal aangeven uit welke richting het kwam.
Onderzoeksresultaten
Om de afhankelijkheid te beoordelen lateralisatie* hersenactiviteit op basis van geluidsintensiteit als reactie op gelabelde neuronen, werden gegevens over de reactiesnelheid van neuronen in de laminaire kern van de kerkuilhersenen gebruikt.
Lateraliteit* - asymmetrie van de linker- en rechterhelft van het lichaam.
Om de afhankelijkheid van de lateralisatie van hersenactiviteit van de reactiesnelheid van bepaalde neuronenpopulaties te beoordelen, werden gegevens over de activiteit van de inferieure colliculus van de hersenen van de resusaap gebruikt, waarna aanvullend de verschillen in de snelheid van neuronen uit verschillende hersenhelften werden berekend. .
Het gemarkeerde lijnmodel van detectorneuronen voorspelt dat naarmate de geluidsintensiteit afneemt, de lateraliteit van de waargenomen bron zal convergeren naar gemiddelde waarden die vergelijkbaar zijn met de verhouding tussen zachte en harde geluiden (1S).
Het hemisferische asymmetriemodel suggereert op zijn beurt dat naarmate de geluidsintensiteit afneemt tot bijna drempelniveaus, de waargenomen lateraliteit naar de middellijn zal verschuiven (1D).
Bij een hogere algehele geluidsintensiteit wordt verwacht dat de lateralisatie intensiteitsinvariant is (inzetstukken in 1S и 1D).
Door te analyseren hoe de geluidsintensiteit de waargenomen richting van het geluid beïnvloedt, kunnen we daarom nauwkeurig de aard bepalen van de processen die op dat moment plaatsvinden: neuronen uit hetzelfde algemene gebied of neuronen uit verschillende hersenhelften.
Het is duidelijk dat het vermogen van een persoon om ITD te onderscheiden kan variëren afhankelijk van de geluidsintensiteit. De wetenschappers zeggen echter dat het moeilijk is om eerdere bevindingen te interpreteren die de gevoeligheid koppelen aan ITD en het oordeel van luisteraars over de richting van de geluidsbron als functie van de geluidsintensiteit. Sommige onderzoeken zeggen dat wanneer de geluidsintensiteit een grensdrempel bereikt, de waargenomen lateraliteit van de bron afneemt. Andere onderzoeken suggereren dat intensiteit helemaal geen effect heeft op de perceptie.
Met andere woorden, wetenschappers laten ‘zachtjes’ doorschemeren dat er in de literatuur weinig informatie bestaat over de relatie tussen ITD, geluidsintensiteit en het bepalen van de richting van de bron. Er zijn theorieën die bestaan als een soort axioma's, algemeen aanvaard door de wetenschappelijke gemeenschap. Daarom werd besloten om alle theorieën, modellen en mogelijke mechanismen van gehoorperceptie in de praktijk in detail te testen.
Het eerste experiment was gebaseerd op een psychofysisch paradigma dat de studie mogelijk maakte van op ITD gebaseerde lateralisatie als functie van de geluidsintensiteit in een groep van tien normaal horende deelnemers.
Afbeelding #2
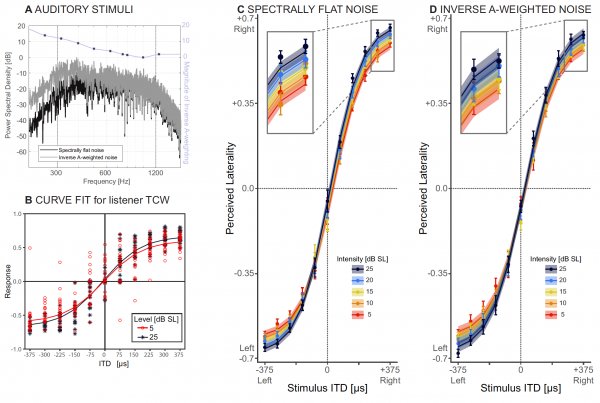
De geluidsbronnen zijn specifiek afgestemd om het grootste deel van het frequentiebereik te dekken waarbinnen mensen ITD kunnen detecteren, d.w.z. van 300 tot 1200 Hz (2А).
Bij elke proef moest de luisteraar de waargenomen lateraliteit aangeven, gemeten als een functie van het sensatieniveau, over een bereik van ITD-waarden van 375 tot 375 ms. Om het effect van de geluidsintensiteit te bepalen, werd een niet-lineair mixed effects model (NMLE) gebruikt dat zowel vaste als willekeurige geluidsintensiteit omvatte.
Plan 2V demonstreert geschatte lateralisatie met spectraal vlakke ruis bij twee geluidsintensiteiten voor een representatieve luisteraar. En het schema 2S toont de onbewerkte gegevens (cirkels) en het aangepaste NMLE-model (lijnen) van alle luisteraars.
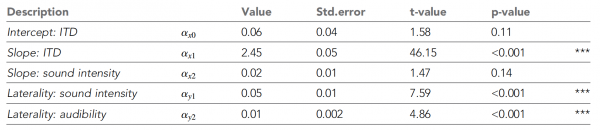
Tabel nr. 1
De bovenstaande tabel toont alle NLME-parameters. Het is duidelijk dat de waargenomen lateraliteit toenam met toenemende ITD, zoals de wetenschappers hadden verwacht. Naarmate de geluidsintensiteit afnam, verschoof de perceptie steeds meer naar de middellijn (inzet in de grafiek). 2C).
Deze trends werden ondersteund door het NLME-model, dat significante effecten van ITD en geluidsintensiteit op de maximale mate van lateraliteit liet zien, wat het model van interhemisferische verschillen ondersteunde.
Bovendien hadden de gemiddelde audiometrische drempels voor zuivere tonen weinig effect op de waargenomen lateraliteit. Maar de geluidsintensiteit had geen significante invloed op de indicatoren van psychometrische functies.
Het belangrijkste doel van het tweede experiment was om te bepalen hoe de resultaten verkregen in het vorige experiment zouden veranderen als rekening werd gehouden met de spectrale kenmerken van de stimuli (geluiden). De noodzaak om te testen op spectraal vlakke ruis bij een lage geluidsintensiteit is dat delen van het spectrum mogelijk niet hoorbaar zijn en dit kan de bepaling van de geluidsrichting beïnvloeden. Bijgevolg kunnen de resultaten van het eerste experiment ten onrechte worden aangezien voor het feit dat de breedte van het hoorbare deel van het spectrum kan afnemen met afnemende geluidsintensiteit.
Daarom werd besloten om nog een experiment uit te voeren, maar dan in omgekeerde volgorde A-gewogen* lawaai
A-weging* toegepast op geluidsniveaus om rekening te houden met de relatieve luidheid die door het menselijk oor wordt waargenomen, aangezien het oor minder gevoelig is voor lage geluidsfrequenties. A-weging wordt geïmplementeerd door rekenkundig een tabel met waarden in octaafbanden toe te voegen aan de gemeten geluidsdrukniveaus in dB.
op de kaart 2D toont onbewerkte gegevens (cirkels) en NMLE-model-aangepaste gegevens (lijnen) van alle deelnemers aan het experiment.
Analyse van de gegevens toonde aan dat wanneer alle delen van het geluid ongeveer even hoorbaar zijn (zowel in de eerste als in de tweede proef), de waargenomen lateraliteit en de helling in de grafiek die de verandering in lateraliteit met ITD verklaart, afnemen met afnemende geluidsintensiteit.
De resultaten van het tweede experiment bevestigden dus de resultaten van het eerste. Dat wil zeggen, in de praktijk is gebleken dat het model dat Jeffress in 1948 voorstelde, niet correct is.
Het blijkt dat de lokalisatie van geluid verslechtert naarmate de geluidsintensiteit afneemt, en Jeffress geloofde dat geluiden door mensen op dezelfde manier worden waargenomen en verwerkt, ongeacht hun intensiteit.
Voor een meer gedetailleerde kennismaking met de nuances van de studie raad ik aan om naar te kijken .
epiloog
Theoretische aannames en praktische experimenten die deze bevestigen, hebben aangetoond dat hersenneuronen bij zoogdieren met verschillende snelheden worden geactiveerd, afhankelijk van de richting van het geluidssignaal. De hersenen vergelijken vervolgens deze snelheden tussen alle neuronen die bij het proces betrokken zijn, om op dynamische wijze een kaart van de geluidsomgeving op te bouwen.
Het model van Jeffresson is eigenlijk niet 100% verkeerd, omdat het gebruikt kan worden om de lokalisatie van de geluidsbron bij kerkuilen perfect te beschrijven. Ja, voor kerkuilen doet de intensiteit van het geluid er niet toe; zij bepalen in ieder geval de positie van de bron. Dit model werkt echter niet bij resusapen, zoals eerdere experimenten hebben aangetoond. Daarom kan dit Jeffresson-model de lokalisatie van geluiden voor alle levende wezens niet beschrijven.
Experimenten met menselijke deelnemers hebben opnieuw bevestigd dat geluidslokalisatie in verschillende organismen anders plaatsvindt. Veel van de deelnemers konden de positie van de bron van geluidssignalen niet correct bepalen vanwege de lage intensiteit van de geluiden.
Wetenschappers zijn van mening dat hun werk bepaalde overeenkomsten vertoont tussen hoe we zien en hoe we horen. Beide processen houden verband met de snelheid van neuronen in verschillende delen van de hersenen, evenals met de beoordeling van dit verschil om zowel de positie te bepalen van de objecten die we in de ruimte zien als de positie van de bron van het geluid dat we horen.
In de toekomst gaan de onderzoekers een reeks experimenten uitvoeren om het verband tussen het menselijk gehoor en zicht gedetailleerder te onderzoeken, waardoor we beter kunnen begrijpen hoe onze hersenen op dynamische wijze een kaart van de wereld om ons heen opbouwen.
Bedankt voor je aandacht, blijf nieuwsgierig en een fijne week allemaal! 🙂
Bedankt dat je bij ons bent gebleven. Vind je onze artikelen leuk? Wil je meer interessante inhoud zien? Steun ons door een bestelling te plaatsen of door vrienden aan te bevelen, , 30% korting voor Habr-gebruikers op een unieke analoog van instapservers, die door ons voor u is uitgevonden: (beschikbaar met RAID1 en RAID10, tot 24 cores en tot 40GB DDR4).
Dell R730xd 2 keer goedkoper? Alleen hier in Nederland! Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - vanaf $99! Lees over
Bron: www.habr.com
